 Tweet
Tweet
Overview
Alternative splicing is a sophisticated cellular process crucial for proteome diversification and significantly impacts cellular development, function, and disease pathways. Given the pivotal role of alternative splicing events, precise mapping and quantification are essential for downstream analyses, notably for associating diseases with specific splicing events. With the rapid evolution of sequencing technologies, specialized tools for particular sequencing methodologies have emerged, allowing for a deeper exploration of splicing intricacies. Traditional short-read RNA-Seq approaches often fall short in capturing full-length isoforms, necessitating computational reconstruction, which can lead to inaccurate assemblies. In contrast, long-read sequencing technologies can sequence isoforms from start to finish in a single read, enabling clear characterization and simultaneous quantification within a single dataset.
What is Alternative Splicing?
Alternative splicing is a regulated cellular mechanism that produces multiple mature mRNA molecules from a single pre-mRNA template. This intricate post-transcriptional process is essential for generating distinct protein isoforms from a single gene, thereby enhancing the proteome’s diversity and complexity. The human genome, with roughly 20,000 protein-coding genes, can produce an extensive array of protein products through alternative splicing.
Mechanism and Regulation of Alternative Splicing
The Complex Mechanism of the Spliceosome
At the heart of the alternative splicing mechanism is the spliceosome, a dynamic macromolecular machine. The spliceosome consists of five small nuclear ribonucleoproteins (snRNPs) and many associated proteins whose main role is to recognize and interact with specific sequences on pre-mRNAs. Notably, these include 5' and 3' splice sites, branch points, and polypyrimidine tracts. Through a complex series of molecular rearrangements, intronic regions are excised and exonic regions are joined together to produce mature mRNA.
Regulation by Splicing Factors
The decision matrix of which exons to include or exclude in the final mRNA is not random. It is orchestrated by splicing factors that either promote or inhibit the action of the splicing machinery. Key players in this regulatory dance include serine/arginine-rich (SR) proteins and heteronuclear ribonucleoproteins (hnRNP).
SR proteins: Usually act as splicing activators. They bind to exonic splicing enhancers (ESEs) and promote spliceosome recruitment to adjacent splice sites.
hnRNP: Often acts as a splicing repressor. Their association with exonic or intronic splicing silencers (ESS or ISS) prevents spliceosome assembly at adjacent splice junctions.
Environmental cues, cellular stress, or developmental stage may cause changes in the levels or activities of these splicing factors, which dictate changes in splicing patterns to suit the cellular environment.
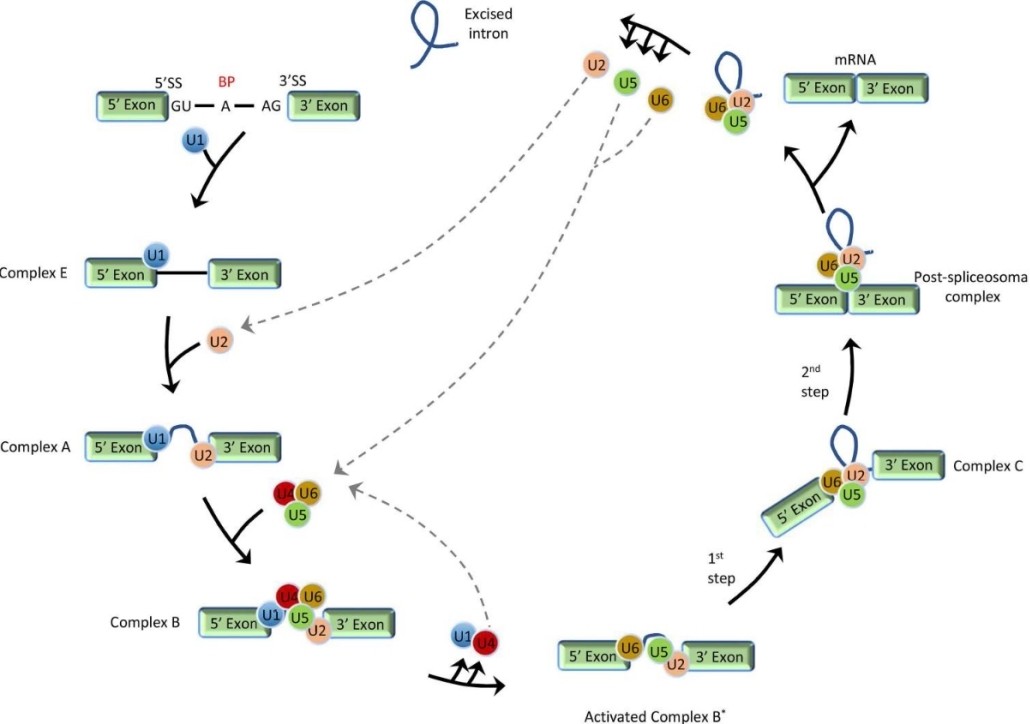
Stepwise schematic presentation of general pre-mRNA splicing. (Jiang, et al., 2021) Human Diseases Associated with Alternative Splicing
Perturbations in the intricate molecular machinery governing alternative splicing can precipitate a spectrum of pathological states. Splicing aberrations, stemming either from mutations directly impinging on splice sites or dysregulation of auxiliary regulatory sequences, are intrinsically linked with a myriad of human pathologies.
Spinal Muscular Atrophy (SMA)
SMA exemplifies the profound consequences of splicing perturbations. The etiological underpinnings of this debilitating neurodegenerative disorder trace back to mutations in the SMN1 gene. However, the clinical trajectory and severity of SMA are modulated by alternative splicing events within its paralog, SMN2. Intriguingly, a mere single nucleotide polymorphism distinguishes SMN1 and SMN2, a discrepancy leading to the differential inclusion of pivotal exons in SMN2. This, in turn, culminates in a truncated protein variant with compromised functionality.
Cancer
The protean nature of alternative splicing is evident in its pervasive role in oncogenesis, shaping tumor biology, aggressiveness, and therapeutic susceptibility. The Bcl-x gene, pivotal in the apoptotic cascade, underscores this with its alternative splicing events birthing two isoforms: the anti-apoptotic Bcl-xL and its pro-apoptotic counterpart, Bcl-xS. A preponderance of Bcl-xL isoforms, resulting from splicing dysregulation, emerges as a recurrent theme across multiple malignancies, invariably fostering tumor resilience and chemoresistance.
Nervous system Disease
The neural milieu isn't immune to splicing aberrations either. A spectrum of neurological afflictions, encompassing both frontotemporal dementia and Parkinson's disease, betray splicing irregularities. Specifically, variations in the splicing patterns of the MAPT gene, encoding tau protein, have been implicated in a cluster of neurodegenerative tauopathies.
Cardiovascular Diseases
The intricate tapestry of cardiac physiology intertwines with alternative splicing. The cardiac troponin T (cTNT) gene, an orchestrator of cardiac muscle contraction, is susceptible to alternative splicing events. Erroneous splicing episodes in cTNT are pathognomonic of dilated cardiomyopathy, exerting detrimental effects on cardiac hemodynamics. Methods for Characterizing Alternative Splicing
RT-PCR and qRT-PCR
Reverse transcription PCR (RT-PCR) and quantitative RT-PCR (qRT-PCR) are well-established methods for validating alternative splicing events identified by high-throughput methods. They can be used to quantify the expression of different splice variants and confirm the presence of specific splice junctions. These methods require careful PCR primer design to ensure specific detection of specific spliced isoforms.
RNA-seq.
RNA-seq has revolutionized transcriptomics by providing a high-throughput approach to the study of transcripts. In the context of alternative splicing, RNA-seq offers the opportunity to detect disease-associated splicing events, even with challenges such as the potential confounding effects of gene expression levels. The fine resolution provided by RNAseq makes it possible to visualize the global dysregulation of splicing and detect mutations in splicing-regulated genes such as SF3B1. Such observations have been documented in a large number of tumors, highlighting the relevance of RNA-seq in uncovering the complex landscape of alternative splicing. As RNA-seq has become more popular for transcriptome studies, various computational tools have been developed to address the challenges of alternative splicing analysis.
Alternative splicing is a sophisticated cellular process crucial for proteome diversification and significantly impacts cellular development, function, and disease pathways. Given the pivotal role of alternative splicing events, precise mapping and quantification are essential for downstream analyses, notably for associating diseases with specific splicing events. With the rapid evolution of sequencing technologies, specialized tools for particular sequencing methodologies have emerged, allowing for a deeper exploration of splicing intricacies. Traditional short-read RNA-Seq approaches often fall short in capturing full-length isoforms, necessitating computational reconstruction, which can lead to inaccurate assemblies. In contrast, long-read sequencing technologies can sequence isoforms from start to finish in a single read, enabling clear characterization and simultaneous quantification within a single dataset.
What is Alternative Splicing?
Alternative splicing is a regulated cellular mechanism that produces multiple mature mRNA molecules from a single pre-mRNA template. This intricate post-transcriptional process is essential for generating distinct protein isoforms from a single gene, thereby enhancing the proteome’s diversity and complexity. The human genome, with roughly 20,000 protein-coding genes, can produce an extensive array of protein products through alternative splicing.
Mechanism and Regulation of Alternative Splicing
The Complex Mechanism of the Spliceosome
At the heart of the alternative splicing mechanism is the spliceosome, a dynamic macromolecular machine. The spliceosome consists of five small nuclear ribonucleoproteins (snRNPs) and many associated proteins whose main role is to recognize and interact with specific sequences on pre-mRNAs. Notably, these include 5' and 3' splice sites, branch points, and polypyrimidine tracts. Through a complex series of molecular rearrangements, intronic regions are excised and exonic regions are joined together to produce mature mRNA.
Regulation by Splicing Factors
The decision matrix of which exons to include or exclude in the final mRNA is not random. It is orchestrated by splicing factors that either promote or inhibit the action of the splicing machinery. Key players in this regulatory dance include serine/arginine-rich (SR) proteins and heteronuclear ribonucleoproteins (hnRNP).
SR proteins: Usually act as splicing activators. They bind to exonic splicing enhancers (ESEs) and promote spliceosome recruitment to adjacent splice sites.
hnRNP: Often acts as a splicing repressor. Their association with exonic or intronic splicing silencers (ESS or ISS) prevents spliceosome assembly at adjacent splice junctions.
Environmental cues, cellular stress, or developmental stage may cause changes in the levels or activities of these splicing factors, which dictate changes in splicing patterns to suit the cellular environment.
Stepwise schematic presentation of general pre-mRNA splicing. (Jiang, et al., 2021) Human Diseases Associated with Alternative Splicing
Perturbations in the intricate molecular machinery governing alternative splicing can precipitate a spectrum of pathological states. Splicing aberrations, stemming either from mutations directly impinging on splice sites or dysregulation of auxiliary regulatory sequences, are intrinsically linked with a myriad of human pathologies.
Spinal Muscular Atrophy (SMA)
SMA exemplifies the profound consequences of splicing perturbations. The etiological underpinnings of this debilitating neurodegenerative disorder trace back to mutations in the SMN1 gene. However, the clinical trajectory and severity of SMA are modulated by alternative splicing events within its paralog, SMN2. Intriguingly, a mere single nucleotide polymorphism distinguishes SMN1 and SMN2, a discrepancy leading to the differential inclusion of pivotal exons in SMN2. This, in turn, culminates in a truncated protein variant with compromised functionality.
Cancer
The protean nature of alternative splicing is evident in its pervasive role in oncogenesis, shaping tumor biology, aggressiveness, and therapeutic susceptibility. The Bcl-x gene, pivotal in the apoptotic cascade, underscores this with its alternative splicing events birthing two isoforms: the anti-apoptotic Bcl-xL and its pro-apoptotic counterpart, Bcl-xS. A preponderance of Bcl-xL isoforms, resulting from splicing dysregulation, emerges as a recurrent theme across multiple malignancies, invariably fostering tumor resilience and chemoresistance.
Nervous system Disease
The neural milieu isn't immune to splicing aberrations either. A spectrum of neurological afflictions, encompassing both frontotemporal dementia and Parkinson's disease, betray splicing irregularities. Specifically, variations in the splicing patterns of the MAPT gene, encoding tau protein, have been implicated in a cluster of neurodegenerative tauopathies.
Cardiovascular Diseases
The intricate tapestry of cardiac physiology intertwines with alternative splicing. The cardiac troponin T (cTNT) gene, an orchestrator of cardiac muscle contraction, is susceptible to alternative splicing events. Erroneous splicing episodes in cTNT are pathognomonic of dilated cardiomyopathy, exerting detrimental effects on cardiac hemodynamics. Methods for Characterizing Alternative Splicing
RT-PCR and qRT-PCR
Reverse transcription PCR (RT-PCR) and quantitative RT-PCR (qRT-PCR) are well-established methods for validating alternative splicing events identified by high-throughput methods. They can be used to quantify the expression of different splice variants and confirm the presence of specific splice junctions. These methods require careful PCR primer design to ensure specific detection of specific spliced isoforms.
RNA-seq.
RNA-seq has revolutionized transcriptomics by providing a high-throughput approach to the study of transcripts. In the context of alternative splicing, RNA-seq offers the opportunity to detect disease-associated splicing events, even with challenges such as the potential confounding effects of gene expression levels. The fine resolution provided by RNAseq makes it possible to visualize the global dysregulation of splicing and detect mutations in splicing-regulated genes such as SF3B1. Such observations have been documented in a large number of tumors, highlighting the relevance of RNA-seq in uncovering the complex landscape of alternative splicing. As RNA-seq has become more popular for transcriptome studies, various computational tools have been developed to address the challenges of alternative splicing analysis.